تمام فعالیتها و صفات موجودات زنده بهوسیله مولکولهای زیستی به نام DNA کنترل میشود. هر DNA از توالی زیرواحدهای نوکلئوتیدی تشکیل شده است. هر نوکلئوتید از یک کربوهیدرات پنجکربنه و چهار گروه بازهای آلی تشکیل میشود. این نوکلئوتید در واحدهای مشخصی به نام ژن کنار هم قرار میگیرند. انواع ژن در سلولهای پروکاریوتی و یوکاریوتی با فرایندی به نام رونویسی به مولکولهای RNA تبدیل میشوند. از رونویسی بعضی ژنها rRNA، بعضی tRNA و بعضی دیگر mRNA تولید میشود. rRNA ساختار ریبوزوم را میسازد. tRNA آمینواسیدهای لازم برای سنتز پروتئین را به ریبوزوم منتقل میکند و mRNA کدهای سنتز پروتئین را ذخیره میکند. ساختار و توالی ژنها در گونههای مختلف متفاوت است. در بیشتر باکتریها ژنها در DNA های حلقوی قرار دارند و بهوسیله توالی مشترکی تنظیم میشوند. اما در بیشتر یوکاریوتها ژن در DNA خطی قرار دارد و با توالیهای مستقل از هم تنظیم میشود. در این مطلب از مجله فرادرس انواع ژن در یوکاریوتها و پروکاریوتها را به همراه مکانیسمهای بیان ژن و تنظیم آن توضیح میدهیم.
ژن چیست؟
ژن واحد انتقال اطلاعات وراثتی است که جایگاه مشخصی از کروموزوم به نام لوکوس را به خود اختصاص میدهد. در سلولهای یوکاریوتی و قارچهای بخش اصلی ژنها در هسته قرار دارند. بخشی از ژنهای جانوران در میتوکندری و بخشی از ژنهای گیاهی در کلروپلاست قرار دارد. در پروکاریوتها اندامکی وجود ندارد. به همین دلیل ژنهای باکتریایی در سیتوپلاسم سلول قرار میگیرد. تعداد و سازمانیافتگی ژنها در گونههای مختلف متفاوت است اما تمام ژنها از زیرواحدهای مشترکی تشکیل شدهاند. ژن بخشی از مولکول DNA است و تمام ژنها از چهار نوکلئوتید آدنین (A)، تیمین (T)، گوانین (G) و سیتوزین (C) تشکیل میشود که بهوسیله پیوندهای فسفودیاستر و پیوند هیدروژنی بین بازها کنار هم قرار میگیرند.
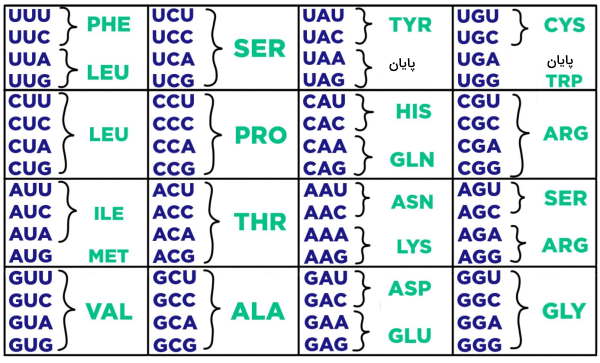
نوکلئوتیدهای ژنهای پروتئینی واحدهای سهتایی به نام کدون میسازند. هر کدون نشاندهنده یک نوع آمینواسید در زنجیره پلیپپتیدی ژن است. کدونها در فرایندی به نام رونویسی بهوسیله آنزیمهای RNA پلیمراز به کدهای RNA و کدهای RNA در فرایند ترجمه به آمینواسیدهای پروتئین تبدیل میشوند. ۶۴ کدون برا ترجمه پروتئینها وجود دارد که در جدول زیر نمایش داده شده است. ۶۱ کدون به آمینواسید ترجمه میشود و ۳ کدون ترجمه پروتئین را پایان میدهد. کدونهای ترجمه آمینواسید در تمام گونهها یکسان است. متیونین و تریپتوفان تنها آمینواسیدهایی هستند که یک کدون دارند.
انواع ژن چیست؟
ژنها را میتوان بر اساس نوع سلول و RNA حاصل از رونویسی به انواع مختلفی تقسیم کرد. در این مطلب ابتدا انواع ژن بر اساس نوع سلول (پروکاریوت و یوکاریوت) و سپس انواع ژن بر اساس RNA را توضیح میدهیم.
انواع ژن بر اساس نوع سلول
ژنها را میتوان بر اساس نوع سلول به انواع پروکاریوتی، یوکاریوتی و ویروسی تقسیمبندی کرد. به مجموع تمام ژنها و بخشهای ساختاری DNA سلولها ژنوم گفته میشود. اندازه ژنوم باکتریها بسیار کوچکتر از ژنوم یوکاریوتها است و ژنوم ویروسی کوچکتر از سلول پروکاریوتی و یوکاریوتی است. تعداد ژنهای پروکاریوتها کمتر از یوکاریوتها و تعداد ژنهای ویروس کمتر از پروکاریوتها است. ژنوم پروکاریوتی و یوکاریوتی از DNA و ژنوم ویروسی از DNA یا RNA تشکیل شده است.
ژن پروکاریوتی
در این سلولها بخش اصلی ژنها روی یک DNA خطی یا حلقوی که کروموزوم باکتری است و بخش کمی از ژنها روی DNA حلقوی یا خطی کوچکتری به نام پلاسمید قرار دارد. پلاسمیدها حاوی ژنهای کمکی هستند، و رشد و بقای باکتری بدون آنها ادامه دارد. به دلیل کوچکتر بودن DNA پروکاریوتی فاصله بین ژنها در این سلولها کمتر است و بعضی از ژنها تنها با یک نوکلئوتید از هم جدا میشوند.
ژنهای باکتری در واحدهای به نام اوپران دستهبندی میشوند. هر اوپران پروتئینهای یک مسیر بیوشیمیایی را کد میکند. تمام ژنهای هر اوپران بهوسیله یک پروموتر رونویسی میشود. ژن پروکاریوتها یکپارچه است و بین آنها توالی غیرکدکننده وجود ندارد. برای مثال اوپران لک یکی از گروههای ژنی باکتری ای. کلی است که آنزیمهای مسیر تجزیه لاکتور به گلوکز و گالاکتوز را کد میکند. ناحیه تنظیمی بخش دیگری از ژنوم باکتریها است. این ناحیه در بالادست و سمت انتهای $$3^prime$$ قرار دارد. پروموتر جایگاه اتصال آنزیمهای RNA پلیمراز برای رونویسی ژنهای اوپران در ناحیه تنظیمی است. به علاوه در این ناحیه جایگاه اتصال به پروتئینهای مهارکننده و فعالکننده ژن وجود دارد. اوپراتور یکی دیگر از جایگاههای تنظیمی ژن باکتریی است که نزدیک اولین ژن اوپران قرار دارد.
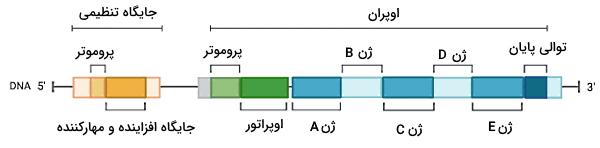
همانندسازی و رونویسی پلاسمیدها جدا از کروموزوم باکتری است. تعداد نوکلئوتیدها و ژنهای پلاسمیدها بسیار کمتر از کروموزوم باکتری است. ژن مقاومت به آنتیبیوتیک و ژن انتقالی، دو ژن اصلی است که در بیشتر پلاسمیدها وجود دارد. ژن انتقالی پروتئینهای لازم برای کانجوگاسیون و ترانسفورماسیون باکتریها را کد میکند. این ژنها کنار یک پروموتور قرار دارند که محل اتصال RNA پلیمراز است. کانجوگاسیون فرایندی است که با ادغام غشا برای مدت کوتاه، پلاسمید از یک باکتری به باکتری دیگر منتقل میشود.
ژن یوکاریوتی
در تعریف انواع ژن اشاره کردیم که بیشتر ژنهای سلولهای یوکاریوتی در هسته و بخشی از آن در اندامکهای میتوکندری و کلروپلاست است. ژنهای هستهای یوکاریوتها روی کروموزومهای خطی قرار دارند. تعداد ژنها در بخشهایی از کروموزوم بیشتر و بخشهایی از آن کمتر است. ژنهای یوکاریوتی بر خلاف پروکاریوتها از توالیهای کدکننده و غیرکدکننده تشکیل میشود. توالیهای غیرکدکننده اینترون و توالیهای کدکننده اگزون نام دارند. اینترونها بین اگزونها قرار میگیرند. قطعات اینترون در تغییرات پس از رونویسی حذف میشوند و در mRNA بالغ وجود ندارد. اگزونها بخشهایی از ژن یوکاریوتی هستند که کدونهای آمینواسید در آنها وجود دارد و پس از حذف اینترونها به هم متصل میشوند. وجود اینترونها به تغییرات تکاملی و نوترکیبی انواع ژن کمک کرده است.
اگزونها و اینترونها در ناحیهای از قطعه ژنی به نام «الگوی خوانش باز» (Open Reading Frame | ORF) قرار دارند. الگوی خوانش باز شامل کدون شروع، کدون پایان و کدونهای بین آنها میشود. بخشی از اولین و آخرین اگزون که در رشته DNA $$5^prime$$ به $$3^prime$$ رونویسی میشود، در mRNA بالغ وجود دارد، اما ترجمه نمیشود. به این دو ناحیه انتهای غیرترجمهای $$3^prime$$ ($$3^prime UTR$$) و $$5^prime$$ ($$5^prime UTR$$) گفته میشود. در بالادست و پاییندست ژنهای پروتئینی نواحی تنظیمی قرار دارد. پروموتر، جایگاه پروتئینهای افزاینده و مهارکننده، و جایگاه پایان در این نواحی قرار دارد.
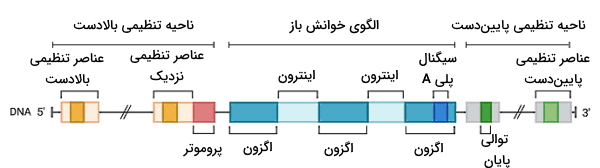
پروموتر نزدیک اولین اگزون قرار دارد. طول این توالی در سلولهای مختلف متفاوت است و سطح بیان ژن را کنترل میکند. هر چه طول پروموتر بیشتر باشد، فضای بیشتری برای اتصال فاکتورهای رونویسی و آنزیم پلیمراز وجود دارد. توالیهای افزاینده و مهارکننده ممکن است نزدیک پروموتر، بین نواحی کدکننده یا در فاصله هزار نوکلئوتیدی انواع ژن قرار داشته باشد.
ژن ویروسی
ویروسها از قطعات DNA یا RNA تکرشتهای یا دورشتهای و پوششی از پروتئینی به نام کپسید تشکیل شدهاند. ژن پروتئینها در ژنوم ویروسها وجود دارد اما بدون ریبوزومها، آنزیمها و مولکولهای زیستی میزبان قادر به رونویسی و ترجمه پروتئینها نیستند. ژنوم بعضی از ویروسها خطی و ژنوم بعضی حلقوی است. ژن بعضی از ویروسها روی یک و ژن بعضی از آنها روی چند قطعه نوکلئیکاسید قرار دارد. ژنوم تکرشته ویروسها ممکن است رشته مثبت یا منفی باشد . رشته مثبت نقش mRNA ویروسی را دارد و با استفاده از سیستم میزبان به پروتئینها ترجمه میشود. رشته منفی ابتدا به رشته مکمل تبدیل و سپس به پروتئین ترجمه میشود.
سازمانیافتگی ژنها در ویروسهای مختلف متفاوت است. برای مثال ژنهای آدونوویروسها مثل ژنهای یوکاریوتی به اگزون و اینترون تقسیم میشود. باکتریوفاژها ویروسهایی هستند که پس از ورود به سلولهای پروکاریوتی تکثیر میشوند. ژنوم باکتریوفاژ T4 مولکول DNA دورشتهای است و پروموتر و توالیهای تنظیمی آن بین نواحی کدکننده ژن قرار دارد. ژن این ویروسها از سه اینترون کدکننده آنزیم تیمیدیلات سنتتاز، ریبونوکلئوتید ردوکتاز هوازی و ریبونوکلئوتید ردوکتاز غیرهوازی تشکیل شده است.
اندازه ژنوم و تعداد ژنهای ویروسی بر اساس نوع میزبان متفاوت است. سرعت تقسیم میتوز در باکتریها بیشتر از سرعت تقسیم سلولهای یوکاریوتی است. باکتریوفاژها برای سازگاری از این سرعت ژنوم کوچک و فشردهای تشکیل میشوند که ژنهای آن با هم همپوشانی دارد. هرپس ویروس و ویروس آبله ویروسهای یوکاریوتی هستند که ژنوم نسبتا بزرگی دارند. ژنوم این ویروسها از ژنهایی تشکیل شده است که آنزیمهای لازم برای همانندسازی ژنوم را کد میکنند. به همین دلیل وابستگی این ویروسها به میزبان کمتر است. ژنوم ویروسهای یوکاریوتی از نواحی ORF تشکیل شده است.
انواع ژن بر اساس RNA
ژنها را میتوان بر اساس محصول رونویسی به انواع mRNA، tRNA و rRNA تقسیم کرد. کدهای سنتز پروتئین در ژنهای mRNA وجود دارد. در بخشهای قبلی این مطلب از مجله فرادرس ساختار ژنهای پروتئینی پروکاریوتها، یوکاریوتها و ویروسها را توضیح دادیم. در این بخش انواع ژن های tRNA و rRNA را بررسی میکنیم. tRNAها و rRNAها نقش مهمی در ترجمه پروتئین دارند. مهمترین تفاوت ژنهای غیرپروتئینی توالیهای تکراری فراوان آنها در ژنوم یوکاریوتی و پروکاریوتی است. به علاوه آنزیمهای RNA پلیمراز متفاوت در رونویسی این ژنها شرکت میکند. RNA پلیمراز I در سلولهای یوکاریوتی ژن rRNA اولیه، RNA پلیمراز II ژن mRNA اولیه و RNA پلیمراز III ژن tRNA، بخشی از rRNA و snRNA (مولکولهای RNA کوچک هسته) را رونویسی میکند.
tRNA آمینواسید را برای ترجمه mRNA به ریبوزوم منتقل میکند. توالی ACC در انتهای ۳ محل اتصال آمینواسید به tRNA است. ساختار دوبعدی تمام tRNAها در گونههای مختلف شبیه برگ شبدر و ساختار سهبعدی آن شبیه حرف L انگلیسی است. این نوکلئیکاسید بهطور میانگین از ۷۶ نوکلئوتید تشکیل میشود. تعدادد ژنهای tRNA در گونههای مختلف متفاوت است. ژنهای tRNA یوکاریوتی در DNA هسته و میتوکندری وجود دارد. ژنهای هستهای انسان روی تمام کروموزومهای به جز کروموزوم ۲۲ و Y قرار دارد.
علاوه بر تعداد ژنها، طول حلقهها و ساقههای ساختار دوم tRNA در گونههای مختلف متفاوت است. اما ساختار مشابهی دارد. هر tRNA از سه حلقه اصلی، یک حلقه متغیر و چهار ساقه تشکیل شده است. در یکی از این حلقهها توالی سهنوکلئوتیدی آنتیکدون وجود دارد که مکمل کدونهای mRNA است و ساقه روبهروی آن به آمینواسید متصل میشود. بعضی از ژن tRNA مثل mRNA یوکاریوتی از توالیهای اگزون و اینترون تشکیل شده است و بعضی از ژنهای tRNA کاملا از هم جدا است.

rRNA در ساختار ریبوزوم وجود دارد و پیوند پپتیدی حین ترجمه را کاتالیز میکند. تعداد زیادی ژن rRNA در ژنوم سلولهای یوکاریوتی و ۱ تا ۱۵ ژن rRNA در ژنوم پروکاریوتها وجود دارد. ژنهای rRNA انسان روی بازوی کوتاه کروموزومهای ۱۳، ۱۴، ۱۵ و ۲۱ قرار دارند. این ژنهای در توالیهای تکراری پشتسرهم قرار گرفتهاند. هر توالی تکراری در پستانداران حدود ۴۳ kb است. پرموتورها، توالیهای تنظیمی و توالیهای پایان بین این تکرارها قرار دارند. پروموتور این ژنها مثل ژنهای mRNA محل شروع رونویسی است و بهوسیله عناصر تنظیمی بالادست کنترل میشود.
از رونویسی ژنهای rRNA بهوسیله RNA پلیمراز I در پستانداران rRNA اولیه ۴۷ S تولید میشود که در فرایند پردازش پس از ترجمه به قطعات ۱۸، ۵٫۸ و ۲۸ S تقسیم میشود. RNA پلیمراز III مولکول RNA ۵S را رونویسی میکند. ژن rRNA پروکاریوتها مثل ژن پروتئینی آنها از یک واحد تنظیمی و یک اوپران تشکیل شده است. rRNA اولیه رونویسی شده از این اوپران حاوی RNAهای ۱۶، ۲۳ و ۵ S است که در پردازش پس از رونویسی از هم جدا میشوند.
بیان انواع ژن
در بخشهای قبلی مطلب انواع ژن توضیح دادیم که ویروسها آنزیمهای لازم برای همانندسازی و رونویسی را ندارند و وابسته به میزبان هستند. در نتیجه با بررسی بیان انواع ژن در سلولهای یوکاریوت و پروکاریوت، بیان ژنهای ویروسی را هم بررسی میکنیم. اولین مرحله بیان ژنها تبدیل دئوکسی ریبونوکلئوتیدهای DNA به ریبونوکلئوتیدهای RNA است. به این فرایند رونویسی گفته میشود. در مرحله بعد بخشهایی از RNA جدا و بعضی گروههای عاملی به آن اضافه میشود. به این فرایند پردازش RNA گفته میشود. در ادامه ریبونوکلئوتیدها با همکاری ریبوزوم و tRNA به توالی آمینواسیدی پروتئین تبدیل میشود. به این فرایند ترجمه گفته میشود. مرحله اول و دوم این فرایند بین تمام ژنها یکسان و مرحله سوم مخصوص ژنهای کدکننده پروتئین است. در این بخش از مطلب مجله فرادرس رونویسی، پردازش و ترجمه انواع ژن را توضیح میدهیم.
رونویسی انواع ژن
رونویسی ژن در سلولهای پروکاریوتی و یوکاروتی مشابه هم است. اما محل رونویسی، تعداد پروتئینهای تنظیمی و نوع آنزیم آنها متفاوت است. ژن سلولهای پروکاریوتی در سیتوپلاسم رونویسی میشود. به همین دلیل رونویسی و ترجمه یک mRNA همزمان انجام میشود. اما در یوکاریوتها ژن در هسته رونویسی و mRNA در سیتوپلاسم بیان میشود.
RNA پلیمراز باکتریایی از پنج زیرواحد تشکیل شده است. دو زیرواحد$$alpha$$، یک زیرواحد $$beta$$ و یک زیرواحد $$beta^prime$$ هسته اصلی آنزیم را میسازند و زیر واحد سیگما جایگاه شروع رونویسی را شناسایی میکند. زیرواحدهای آلفا به کنار هم قرار گرفتن زیرواحدهای آنزیم روی DNA کمک میکند. زیرواحد $$beta$$ به ریبونوکلئوتیدهایی که رشته جدید RNA را میسازند و زیرواحد $$beta^prime$$ به رشته DNA الگو متصل میشود. جهت رشته DNA الگو از $$3^prime$$ به $$5^prime$$ است. به اولین نوکلئوتید جایگاه شروع رونویسی ناحیه ۱+، نوکلئوتیدهای قبل از آن را با علامت منفی و نوکلئوتیدهای بعد از آن را با علامت مثبت نشان میدهند. رشته غیرالگو همجهت RNA در حال سنتز است. نواحی بالادست و پایین دست ژن را بر اساس این رشته تعیین میکنیم. توالی سمت$$5^prime$$ مثبت و توالی سمت $$3^prime$$ منفی است.
پروموتر جایگاه اتصال آنزیم پلیمراز برای شروع رونویسی است. اگرچه توالی این جایگاه در گونههای مختلف پروکاریوت متفاوت است، دو بخش کاملا یکسان ۱۰- (TATAAT) و ۳۵- (TTGACA) در همه پروموترها وجود دارد. پس از اتصال زیرواحد سیگما به این نواحی، بقیه زیرواحدهای RNA پلیمراز به DNA متصل میشود. مرحله شروع با رونویسی ۱۰ نوکلئوتید اول تمام میشود. مرحله طویلسازی رونوسی با جدا شدن زیرواحد سیگما شروع میشود. با جدا شدن این زیرواحد پلیمراز در طول DNA حرکت، پیوند هیدروژنی بین بازها را هیدرولیز و رشته RNA را در جهت $$5^prime$$ به $$3^prime$$ رونویسی میکند. بر اساس نوع ژن رونویسی با دو روش وابسته به پروتئین Rho و مستقل از این پروتئین پایان مییابد.
- روش وابسته به پروتئین Rho: در این روش پروتئین Rho پشت سر آنزم پلیمراز حرکت میکند. در انتهای رونویسی توالی از نوکلئوتیدها G وجود دارد که سبب توقف آنزیم پلیمراز میشود. در این شرایط پروتئین Rho با آنزیم برهمکنش میدهد و mRNA تازه سنتز شده از آنزیم جدا میشود.
- روش مستقل از پروتئین Rho: در این روش توالی ویژه DNA الگو رونویسی را پایان میدهد. نزدیک انتهای رونویسی توالی از نوکلئوتیدهای GC رونویسی میشود. RNA تا میخورد و با پیوند هیدروژنی بین بازها ساختار سنجاقسری پایداری تشکیل میشود. این ساختار از حرکت بیشتر آنزیم پلیمراز جلوگیری میکند. انتهای ژن از توالی AT تشکیل شده که پیوندهیدروژنی آنها ناپایدار است و به راحتی شکسته میشود. در نتیجه آنزیم پلیمراز از DNA و RNA از آنزیم جدا میشود.
برخلاف RNA پلیمراز پروکاریوتی، پلیمرازهای یوکاریوتی به فاکتورهای رونویسی برای اتصال به DNA نیاز دارند. RNA پلیمراز I و ژنهای rRNA در هستک قرار دارند. RNA پلیمراز II و III در هسته قرار دارند. پروموتر RNA پلیمراز II یوکاریوتها از پروکاریوتها بلندتر است و توالی غنی از AT آن (TATAAA) در ناحیه ۳۰- قرار دارد. در پروموتر RNA پلیمراز I دو ناحیه غنی GC در ناحیه ۴۵- و ۲۰+ وجود دارد. در این ژنها دو ناحیه ۱۸۰- و ۱۰۵- پروموترهای کمکی RNA پلیمراز I هستند که به افزایش بیان ژن rRNA کمک میکنند. پروموتر RNA پلیمراز III در بالادست یا بین توالیهای ژنی است.
فاکتورهای رونویسی با نماد TFII و حروف A تا J انگلیسی نمایش داده میشود. این فاکتورهای پروتئینها بازی هستند که کمپلکس شروع رونویسی را تشکیل میدهند. این کمپلکش از ۴۰ پروتئین تشکیل شده است. شروع رونویسی tRNA و rRNA به فاکتورهای کمتری نیاز دارد. TFH آنزیمی است که فعالیت هلیکاری و فسفاتازی دارد. این آنزیم پیوند هیدروژنی بین نوکلئوتیدها را هیدرولیز به انتهای کربوکسیلی آنزیم RNA پلیمراز II گروه فسفات اضافه میکند. با اضافه شدن گروه فسفات کنفورماسیون پلیمراز تغییر میکند و از فاکتورهای رونویسی جدا میشود.

وجود پروتئینهای هیستون& یکی دیگر از تفاوتهای دیگر رونویسی پروکاریوت و یوکاریوت است. قبل از شروع رونویسی این پروتئینها باید از DNA جدا شوند. این فرایند بهوسیله کمپلکس پروتئینی FACT انجام میشود. این پروتئین همزمان با حرکت RNA پلیمراز پروتئینهای هیستون را جدا میکند و پس رونویسی DNA را به حالت اول برمیگرداند. RNA یوکاریوتها مثل پروکاریوتهادر جهت $$5^prime$$ به $$3^prime$$ رونویسی میشود. پایان رونویسی RNA پلیمرازهای یوکاریوتی متفاوت است. RNA پلیمراز II از چند تا چند هزار باز را رونویسی میکند و توالی پایان ندارد. قبل از پایان رونویسی بخشی از mRNA شکسته و جدا میشود. به محل شکست جایگاه پایان رونویسی گفته میشود. آنزیمهای $$5^prime$$ اگزونوکلئاز باقیمانده mRNA را هیدرولیز میکند. در انتهای ژنهای rRNA توالی ۱۸ نوکلئوتیدی پایان رونویسی وجود دارد که بهوسیله پروتئینهای خاتمه شناسایی و رونویسی تمام میشود. در پایان رونویسی ژنهای tRNA ساختار سنجاقسری مثل پایان رونویسی پروکاریوتها تشکیل میشود.
پردازش RNA
پردازش مجموعهای از واکنشها است که در سلولهای پروکاریوتی و یوکاریوتی RNA اولیه را به RNA بالغ تبدیل میکند. تفاوت پردازش mRNA بین یوکاریوتها و پروکاریوتها بیشتر است. در پروکاریوتها mRNA همزمان با رونویسی ترجمه میشود. در یوکاریوتها قبل از خروج mRNA از هسته گروههای عاملی به دو سر مولکول اضافه و اینترونها در فرایند «پیرایش» (Splicing) از mRNA خارج میشود.
- اضافه شدن گروههای عاملی: اضافه شدن گروههای عاملی به خروج mRNA از هسته کمک و از این مولکول در برابر آنزیمهای نوکلئاز محافظت میکند. کلاهک $$5^prime$$، نوکلئوتید گوانین متصل به متیل است که به انتهای $$5^prime$$ اضافه میشود و به اتصال ریبوزوم و mRNA کمک میکند. دم پلی A توالی از نوکلئوتیدهای آدنین در انتهای $$3^prime$$ مولکول mRNA است. وقتی RNA پلیمراز در رونویسی به سیگنال پلی A میرسد، mRNA بهوسیله آنزیمهای نوکلئاز شکسته میشود و آنزیم دیگری توالی ۱۰۰ تا ۲۰۰ نوکلئوتیدی آدنین را به mRNA اضافه میکند. این توالی به خروج mRNA از هسته کمک میکند.
- پیرایش mRNA: در فرایند پیرایش بهوسیله کمپلکس اسپلایسوزوم از mRNA جدا میشود. این کمپلکس از پروتئین و snRNA تشکیل شده است. این کمپلکس اینترونها را با توالی ابتدا، انتها و میانی شناسایی میکند. ابتدای اینترون (سمت $$5^prime$$) توالی GU، انتهای آن (سمت $$3^prime$$) توالی AG و در میانه نوکلئوتید A وجود دارد. در این فرایند ابتدا پیوند انتهای $$5^prime$$ اینترون شکسته میشود. انتهای $$5^prime$$ به نوکلئوتید A متصل و حلقه بستهای تشکیل میشود. در مرحله بعدی پیوند اتنهای $$3^prime$$ شکسته، دو اگزون به هم متصل و اینترون خارج میشود.
rRNA پروکاریوتی و یوکاریوتی به شکل یک RNA اولیه رونویسی و سپس به زیرواحدهای ریبوزوم شکسته میشود. tRNA پروکاریوتی مثل mRNA و rRNA به شکل یک RNA اولیه رونویسی و پس از پردازش به tRNA های مختلف تبدیل میشود. tRNA در پنج مرحله پردازش میشود. tRNA یوکاریوتی در هسته پردازش و در سیتوپلاسم به آمینواسید متصل میشود.
- انتهای $$5^prime$$ مولکول tRNA اولیه جدا میشود.
- انتهای $$3^prime$$ مولکول tRNA اولیه جدا میشود.
- در تمام یوکاریوتها، بعضی از باکتریها و بعضی از آرکئاباکترها توالی CCA به انتهای $$3^prime$$ اضافه میشود. در بعضی از باکتریها و آرکیباکترها توالی CCA در انتهای $$3^prime$$ وجود دارد.
- ساختار بعضی از نوکلئوتیدها با تغییر باز نیتروژنی تغییر میکند. تبدیل آدنین به سودویوریدین یا اینوزین و تبدیل یوریدین دیهیدرویوریدین متداولترین تغییرات نوکلئوتیدی tRNA است.
- در یوکاریوت ها و پروکاریوتها اینترونها جدا میشوند. این قطعات در پروکاریوتها وجود ندارد.

ترجمه انواع ژن
ژن یوکاریوتی و پروکاریوتی با فرایندهای مشابهی ترجمه میشود. در این بخش از مطلب مجله فرادرس ترجمه انواع ژن در پروکاریوتها و سپس تفاوت آن با یوکاریوتها را توضیح میدهیم. اولین مرحله ترجمه ژن در پروکاریوتها تشکیل کمپلکس شروع است. این کمپلکس از زیرواحد کوچک ریبوزوم (۳۰ S)، mRNA، سه فاکتور رونویسی و فرمیل-متیونیل tRNA تشکیل شده است. در این سلولها تشکیل پیوندی هیدروژنی بین توالی غنی از نوکلئوتید G (شاین دالگارنو | AGGAGG) و rRNA ریبوزوم را به mRNA متصل میکند. در تمام مراحل ترجمه انرژی تغییر کنفورماسیون پروتئینها و حرکت ریبوزوم با هیدرولیز GTP تامین میشود. در مرحله بعد آنتیکدون tRNA آغازگر با کدون AUG در mRNA پیوند هیدروژنی تشکیل میدهد. پس از اتصال tRNA آغازگر زیرواحد بزرگتر (۵۰ S) به زیرواحد کوچکتر متصل و ریبوزوم تشکیل میشود.
در مرحله اول شروع ترجمه ژن سلولهای یوکاریوتی،زیرواحد ۴۰ S ریبوزوم به کلاهک $$5^prime$$ mRNA متصل میشود و متیونین tRNA آغازگر به فرمیل متصل نیست. ریبوزوم برای پیدا کردن کدون AUG روی mRNA حرکت میکند. در بعضی از سلولها ترجمه از اولین کدون AUG شروع میشود و در بعضی از سلولها ترجمه از کردون AUG که نزدیک توالی $$5′-GCCRCCAUGG-3′$$ شروع میشود. پس از پیدا شدن AUG آغازگر زیرواحد ۶۰ S به ۴۰ S متصل و طویلسازی زنجیره پلیپپتیدی شروع میشود.
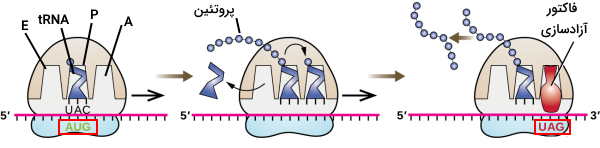
ریبوزوم از سه جایگاه A، P و E برای اتصال به tRNA تشکیل شده است. جایگاه A محل ورود tRNA متصل به آمینواسید، زیرواحد P محل قرار گرفتن tRNA متصل به زنجیره پلیپپتیدی و زیرواحد E محل خروج tRNA بدون آمینواسید است. فقط tRNA آغازگر به جای جایگاه A وارد جایگاه P میشود. با هر بار حرکت ریبوزوم در مرحله طویلسازی tRNA جایگاه A وارد جایگاه P و tRNA جایگاه P وارد جایگاه E میشود. در این حرکت به کمک فعالیت آنزیمی rRNA بین آمینواسید جدید و رشته پروتئینی تازه سنتز شده پیوند پپتیدی برقرار میشود. این مرحله در پروکاریوتها و یوکاریوتها یکسان است.
ترجمه ژن زمانی پایان مییابد که یکی از کدونهای پایان (UAA، UAG و UGA) در جایگاه A قرار بگیرد. فاکتورهای پروتئینی آزادسازی این کدونها را شناسایی میکنند. در این مرحله rRNA پیوند بین اولین آمینواسید و آخرین tRNA را هیدرولیز میکند و پلیپپتید سنتز شده از ریبوزوم جدا میشود. دو زیرواحد ریبوزوم از هم و از mRNA جدا میشود و ترجمه پایان مییابد.
تنظیم بیان انواع ژن
زمان، نوع بافت و فراوانی بیان ژن بهوسیله پروتئینهای تنظیمی و تغییرات شیمیایی DNA تنظیم میشود. به همین دلیل مجموعه ژنهای تمام سلولهای انسان یکسان است. اما ژن رنگ چشم در سلولهای عصبی بیان نمیشود. تنظیم بیان ژن مکانیسم اصلی تمایز سلولها و پاسخ به تغییرات محیطی است. بیان ژن سلولهای پروکاریوتی با اتصال پروتئینهای فعالکننده، مهارکننده و القاکننده به اوپراتور تنظیم میشود. پروتئینهای فعالکنندهها، بیان ژن را افزایش میدهند. پروتئینهای مهارکنندهها، بین ژن را متوقف میکنند و مولکولهای القاکنندهها، پروتئینهای مهارکننده را غیرفعال میکنند.
برای مثال ای. کلی برای رشد و بقا به آمینواسید تریپتوفان نیاز دارد. این آمینواسید بهوسیله ژنهای باکتری سنتز یا از محیط دریافت میشود. در شرایطی که غلظت تریپتوفان محیطی زیاد است، اتصال این آمینواسید به پروتئینهای مهارکننده اوپران trp منجر به تغییر شکل پروتئین و اتصال آن به اوپراتور میشود. اتصال مهارکننده-اوپراتور حرکت آنزیم پلیمراز را مهار میکند. در نتیجه سنتز تریپتوفان در سلول کاهش مییابد.
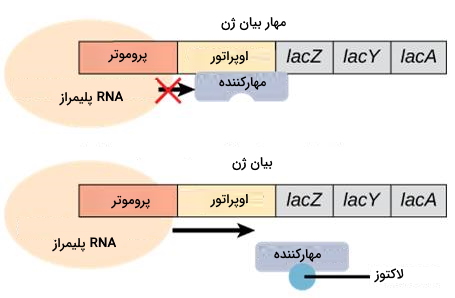
اوپران لَک (lac) یکی دیگر از ژنهای متابولیکی ای. کلی است. زمانی که غلظت گلوکز محیط کم است، ای. کلی از لاکتور به عنوان منبع انرژی استفاده میکند. اوپران lac ژن آنزیمهای تجزیه لاکتوز را کد میکند. اگر غلظت گلوکز محیط کافی باشد، اتصال پروتئینهای مهارکننده به اوپراتور lac از بیان ژنها جلوگیری میکند. در کمبود گلوکز، مولکولهای لاکتوز به مهارکننده متصل و مهارکننده از DNA جدا میشود. در ادامه RNA پلیمراز ژن را رونویسی میکند. لاکتوز، مولکول القاکننده این اوپران است. اما اگر فقط لاکتوز در محیط وجود داشته باشد، باکتری به بیان بیشتر آنزیمهای تجزیه این کربوهیدرات نیاز دارد. در این شرایط اتصال cAMP به پروتئین فعالکننده متابولیکی با تغییر کنفورماسیون این پروتئین و اتصال به پروموتر همراه است. پروتئین فعالکننده اتصال پلیمراز به پروموتر و بیان ژن را افزایش میدهد.
تنظیم بیان ژن در یوکاریوتها پیچیدهتر از پروکاریوتها است و در سطح DNA، mRNA و پروتئین انجام میشود. اولین سطح تنظیم بیان ژن یوکاریوتها تغییرات اپیژنتیکی مولکول DNA و پروتئینهای هیستون است. در این تغییرات گروههای عاملی مختلف به نوکلئوتیدهای DNA یا آمینواسیدهای هیستون اضافه میشود. این گروهها برهمکنش DNA-پروتئین و بیان زن را تغییر میدهد. برای مثال اضافه شدن گروههای متیل به هیستون با افزایش برهمکنش DNA-هیستون، فشرده شدن DNA و کاهش بیان ژن همراه است. اما اضافه شدن گروههای استیل، کاهش برهمکنش، باز شدن DNA و افزایش بیان ژن را به دنبال دارد.
اتصال پروتئینهای تنظیمی به پروموتر، توالی افزاینده و توالی مهارکننده یکی دیگر از مکانیسمهای تنظیم بیان ژن در یوکاریوتها است. اتصال فاکتورهای رونویسی به پروموتر شروع، طویلسازی و پایان رونویسی را تنظیم میکند. اتصال پروتئینها به توالی مهارکننده با مهار بیان ژن و به توالی افزاینده با افزایش بیان ژن همراه است. تغییر الگوی اتصال اگزونها در مرحله پیرایش یکی دیگر از مکانیسمهای تنظیم بیان ژن یوکاریوتی است. برای مثال ژنی با چهار اگزون را تصور کنید. با اتصال اگزونهای ۱، ۲ و ۳ در پیرایش mRNA یک نوع پروتئین و با اتصال اگزونهای ۱، ۳ و ۴ پروتئین دیگری سنتز میشود.
کنترل پایداری mRNA مکانیسم دیگر تنظیم بیان ژن یوکاریوتی است. هر چه زمان حضور mRNA در سیتوپلاسم بیشتر باشد، ریبوزومها پروتئین بیشتری سنتز میکنند. پروتئینهای اتصالی به RNA، کلاهک انتهای $$5^prime$$ و دم پلی A از تجزیه mRNA بهوسیله آنزیمهای نوکلئاز جلوگیری میکنند و بیان ژن را افزایش میدهند. اتصال miRNA همراه کمپلکس پروتئینی به mRNA، با تجزیه mRNA و کاهش بیان ژن همراه است.

در مرحله سنتز پروتئین ترجمه توالی مشخصی از mRNA ممکن است سیگنال پیام تعیینکننده محل پروتئین یا هیدرولیز این مولکول باشد. برای مثال ترجمه توالی مشخصی از mRNA پروتئینهای شبکه اندوپلاسمی با توقف ترجمه در سیتوپلاسم تا زمان اتصال ریبوزومها به این اندامک همراه است. تغییرات پس از ترجمه آخرین مرحله تنظیم بیان زن یوکاریوتی است. در این مرحله اضافه شدن برگشتپذیر یا برگشتناپذیر گروههای عاملی مختلف عملکرد پروتئین را تغییر میدهد. در این مرحله پروتئینهایی که ساختار چهارم درستی ندارند، بهوسیله مولکولهای یوبیکوئیتین نشانهگذاری و بهوسیله آنزیمهای پروتئاز تجزیه میشوند.
انواع جهش ژن
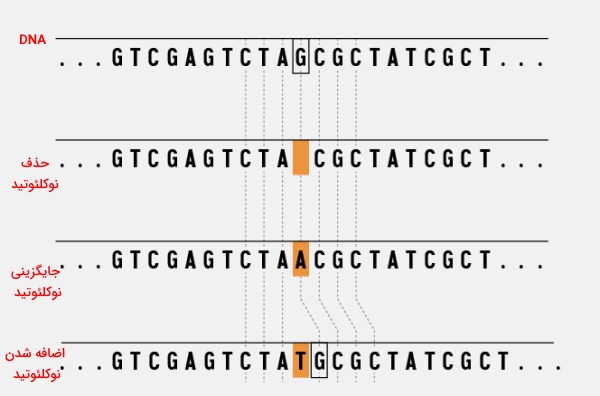
تغییر توالی نوکلئوتیدهای انواع ژن، جهش نام دارد. این تغییر ممکن است به دلیل خطا در همانندسازی DNA (جهش خودبهخودی)، آلودگی ویروسی یا موتاژنها (جهش القایی) ایجاد شود. تغییراتی که در سلولهای زاینده غدد جنسی یا گامتها ایجاد میشود به نسل بعدی منتقل میشود. اما تغییراتی که در سلولهای دیگر ایجاد میشود، ممکن است با بروز بیماری همراه شود. در این تغییرات ممکن است ناحیه کوچک یا بزرگی از ژن حذف یا اضافه شود یا با نوکلئوتیدهای دیگری جایگزین شود.
حذف یا اضافه سه نوکلئوتید همزمان با هم با حذف یا اضافه شدن یک آمینواسید از زنجیره پلیپپتیدی نهایی همراه است. در این حالت پروتئین ممکن است عملکرد خود را داشته باشد، عملکرد آن کاهش پیدا کند یا کاملا غیر عملکردی شود. اضافه شدن یا حذف شدن یک نوکلئوتید الگوی خوانش باز ژن را تغییر میدهد و پروتئین حاصل از ترجمه غیرعملکردی خواهد بود. جایگزینی یک نوکلئوتید با نوکلئوتید دیگر ممکن است بیاثر باشد یا جهش ایجاد کند. اگر با جایگزینی نوکلئوتید کدون به کدون دیگر همان آمینواسید تبدیل شود، جهش بیاثر است. اما اگر کدون به کدون پایان یا آمینواسید دیگر تبدیل شود، پروتئین حاصل از ترجمه غیرعملکردی خواهد بود.
سوالات متداول
در این بخش از مطلب مجله فرادرس به تعدادی از سوالات متدوال پیرامون انوع ژن پاسخ میدهیم.
الل چیست؟
الل یکی از دو یا چند شکل توالی DNA در لوکوس ژنی است. هر فرد دیپلوئید، از هر والد یک الل دریافت میکند. اگر دو الل شبیه هم باشند، فرد برای آن ژن هموزیگوت و اگر دو الل متفاوت باشند، فرد برای آن الل هتروزیگوت است. رابطه بین اللها در ژنهای مختلف، متفاوت است. اللها ممکن است نسبت به هم غالب، مغلوب، غالب ناقص یا همبارز باشند. در رابطه غالب و مغلوب یک الل غالب برای بروز صفت کافی است. اما برای بروز صفت مغلوب، فرد باید برای الل مغلوب هموزیگوت باشد. در رابطه غالب ناقص هر دو الل به اندازه یکسان بروز میکنند و صفت ایجاد شده ترکیبی از دو الل است. در رابطه همبارزی در فرد هتروزیگوت همزمان صفت هر دو الل را به طور کامل نشان میدهد.
ژنوتیپ چیست؟
ژنوتیپ جایگاه ژنها در کل ماده ژنتیکی فرد را نشان میدهد. به کمک ژنوتیپ میتوان اللهای مختلف یک ژن را مشخص کرد. برای نمایش ژنوتیپ میتوان از نمادها یا توالی بازهای DNA استفاده کرد.
فنوتیپ چیست؟
فنوتیپ صفتی است که از بروز هر ژن ایجاد میشود. ژنوتیپ و عوامل محیطی بروز این صفات را کنترل میکنند. قد، رنگ چشم، نوع گروه خونی، رنگ مو و بعضی از بیماریها ازجمله فنوتیپهای انسان هستند.
خزانه ژنی چیست؟
خزانه ژنی مجموع ژنهای یک جمعیت در یک زمان مشخص است. از این اصطلاح برای ژنهای یک گونه استفاده میشود. جهش، انتخاب طبیعی و رانش ژن، ترکیب خزانه ژنی را تغییر میدهد. تکامل و سازگاری یک جمعیت به تنوع و اندازه خزانه ژنی بستگی دارد. هر چه خزانه ژنی بزرگتر و متنوعتر باشد، سازگاری جمعیت با تغییرات محیطی بیشتر است.
رانش ژن چیست؟
رانش ژن یکی از مکانیسمهای تکامل و تغییر فراوانی ژنها در یک جمعیت، به دلیل تغییرات تصادفی است. این تغییرات ممکن است منجر به حذف کامل ژن در جمعیت و کاهش تنوع شود. اثر رانش ژن در جمعیتهای بزرگتر، کمتر است. در این فرایند برخلاف انتخاب طبیعی، اللها برای نگهداشتن صفات سازگارتر با محیط، حذف نمیشوند. به همین دلیل ممکن است صفت سازگارتر حذف شود و و فراوانی صفتی که کارایی کمتری دارد افزایش یابد. زلزله، سیل و آتشسوزی ازجمله عوامل محیطی رانش ژن است.
جریان ژنی چیست؟
جریان یا شارش ژن یکی از مکانیسمهای مهم ایجاد تنوع در گونههای یکسان است. این فرایند به دلیل مهاجرت افراد بین جمعیت گونههای مشابه ایجاد میشود. این فرایند فراوانی اللها در کل یک گونه را تغییر نمی دهد. اما فراوانی اللها در زیستگاههای مختلف گونه تغییر میکند.
source